東京医科大学免疫学分野の横須賀忠主任教授ら研究チーム「免疫チェックポイント分子LAG-3による新たな免疫抑制メカニズムを発見」~ チェックポイント阻害剤抗LAG-3抗体によるがん免疫療法の理解に期待 ~
・免疫チェックポイント分子LAG-3と、その相手方であるMHCクラスIIとが結合した分子の集合体を、世界で初めて可視化しました。
・T細胞が、LAG-3を介した『トランスエンドサイトーシス』というひきちぎりの機序により、抗原提示細胞からMHCクラスII分子を奪い取る、全く新しい免疫調節機構を発見しました。
・LAG-3によってMHCクラスIIが奪い取られた抗原提示細胞は、T細胞の活性化ができなくなるだけでなく、病原性CD4陽性T細胞による大腸炎の進行を抑えられる可能性も示されました。
【研究の背景】
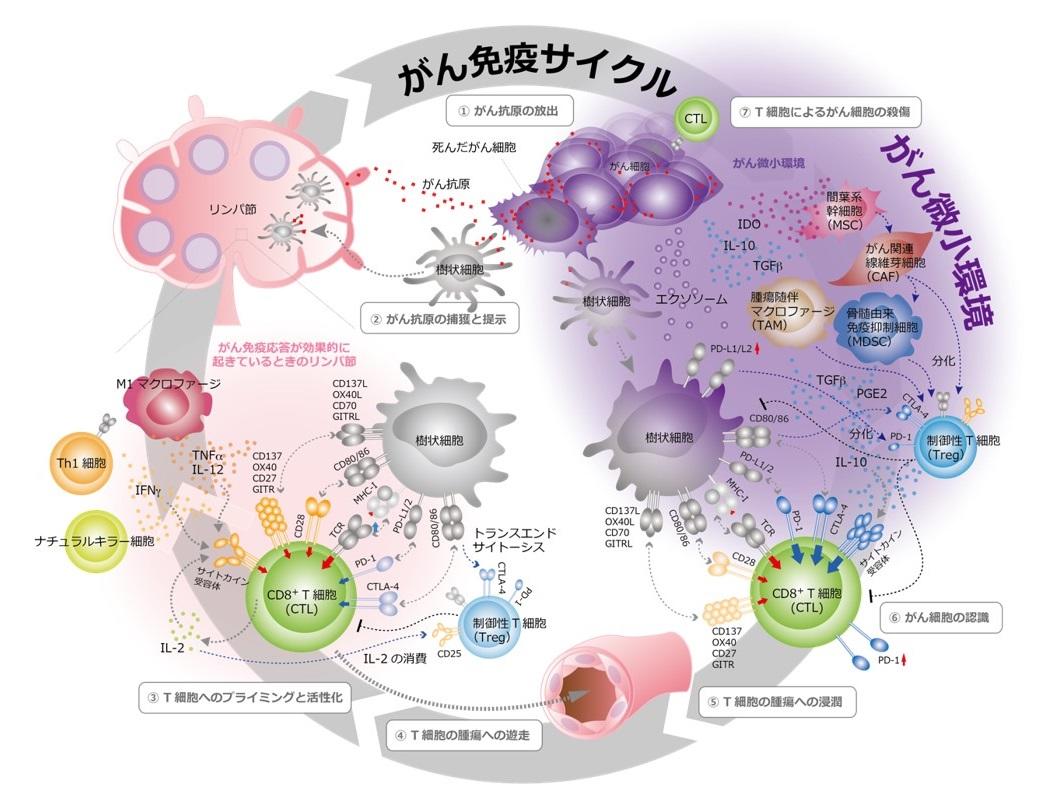
私達の体内では1日に3000個のがん細胞が生まれていますが、発がんには至りません。「免疫監視」と呼ばれるチェック機構が働き、免疫細胞が小さながんの芽を摘んでいるからです。がんに近い所属リンパ節では、がんの抗原が免疫細胞、特にT細胞へと提示され、活性化したT細胞は体内を循環し、がん細胞に遭遇するとこれを殺します。この好循環は、「がん免疫サイクル」と呼ばれています(図1)。しかし、ひとたび体力が衰えたり、がんが少し大きくなったりすると、がんは周囲を自らにとって住みやすい環境に作り変える、いわゆる「がん微小環境」を構築します。このようにがん免疫サイクルが破綻することで、がん細胞は急速に成長し発がんに至ります。
本研究室はこれまで、抗原提示細胞やがん細胞の細胞膜を模倣した人工平面脂質膜(SLB)と超解像顕微鏡とを用いることで、1分子レベルの解析も可能な独創的かつ先端的な分子イメージングシステムを構築してきました(Yokosuka et al,, Nat Immunol (2005))(図2)。
このシステムを用いて、マウスPD-1とPD-L1とが結合してできるPD-1の集合体「PD-1マイクロクラスター」がT細胞受容体(TCR)シグナルを抑えT細胞を疲弊状態に陥らせている現象を解明し、高い評価を得ています(Yokosuka et al., J Exp Med (2012)、Takehara et al., Commun Biol (2021)、Nishi et al., Nat Commun (2023))。本研究では、この分子イメージングシステムをLAG-3観察用にバージョンアップし、LAG-3を介したT細胞抑制機構の解明を目指しました。
【本研究で得られた結果・知見】
今回、T細胞上のLAG-3の動きを解析するため、LAG-3の1細胞分子イメージングの新たな実験系を確立しました。まず、LAG-3のリガンドであるMHCクラスII(MHC-II)を組み入れた「人工がん細胞脂質二重膜」を作成し、その上にLAG-3を発現しているT細胞をのせ、LAG-3の挙動を観察しました。T細胞は数分をかけて脂質二重膜に接着し、MHC-IIとの結合を機にLAG-3分子が数十個集まったクラスターが形成され、その後、LAG-3クラスターは真ん中に集まっていくのが観察されました(図3)。
抗LAG-3抗体が一昨年に難治性のがん患者への治療として米国で承認され、今後、日本でも現在使用されている免疫チェックポイント分子阻害抗体(ICB)との併用療法が現実的になってきました。しかし、LAG-3自身の免疫抑制作用が理解されない状況は抗LAG-3抗体を投与する患者の選定や、複合免疫療法などの可能性を考慮する際の科学的な根拠が欠けているだけでなく、抗体投与によって発生する副作用の予測も十分には行えないことが予想されます。これまで、がん免疫において抗LAG-3抗体はCD8 T細胞を活性化することでがんの縮小に繋がると考えられてきましたが、本研究結果から、CD8 T細胞だけでなくCD4 T細胞の活性化も誘導できる可能性が示されました。これらの結果は、抗PD-1抗体およびそれ以外のICBや化学療法を併用する際の選定の一助となり、またCD4 T細胞による炎症が副作用として発生する可能性を事前に考慮する根拠となることが予想されます。それにより、今後、抗LAG-3抗体をがん患者に使用する際に、より効率的かつ安全性の高い投与法の開発に繋がることが期待されます。
【語句説明】
*トランスエンドサイトーシス
一般に、ある細胞が受容体などを介して、相手方の細胞の一部を含めてかじり取る機構を示す。貪食様式の1つであり、トロゴサイトーシスとも呼ばれる。赤痢アメーバが腸管上皮をかじり取り、病状の悪化に繋がる現象がよく知られており、単細胞生物から哺乳動物まで進化的に広く保存されている。免疫細胞でも頻繁に使われ、奪い取った分子を自分の細胞表面に発現し新たな機能を獲得したり、制御性T細胞が免疫チェックポイント分子CTLA-4を介して抗原提示細胞からリガンドCD80/CD86を奪い取り、免疫機能が低下したりする例が知られている。
【掲載誌名・DOI】
掲載誌名:Cell reports
DOI:10.1016/j.celrep.2024.114655
【論文タイトル】
Indirect suppression of CD4 T-cell activation through LAG-3-mediated trans-endocytosis of MHC class II
【著者】
Ei Wakamatsu*, Hiroaki Machiyama, Hiroko Toyota, Arata Takeuchi, Ryuji Hashimoto, Haruo Kozono, Tadashi Yokosuka*
(*責任著者)
【主な競争的研究資金】
本研究は、文部科学省基盤研究(JP25113725, JP15H01194, JP16H06501, JP17H03600, JP19K07718, JP19K22545, JP20H03536, JP23H02775)、さきがけ慢性炎症(U1114011)、新学術領域「ネオ・セルフ」(JP16H06501)、学術変革A「自己指向性免疫」(JP23H04790)、「私立大学戦略的研究基盤形成支援事業」(S1511011)、内藤記念科学振興財団、武田科学振興財団の支援を受けています。
本件に関するお問い合わせ先
企画部 広報・社会連携推進室
- 住所
- 〒160-8402 東京都新宿区新宿6-1-1
- TEL
- 03-3351-6141
- d-koho@tokyo-med.ac.jp