【名古屋大学】栄養条件に応じ単細胞-多細胞をスイッチする遺伝子特定~多細胞生物の出現や進化の仕組み理解に寄与~
【本研究のポイント】
・海洋由来の黒色酵母注1)株が栄養条件に応じて単細胞性増殖と多細胞体形成を切り換えることを発見した。
・単細胞性―多細胞性の切り換えに必要な10遺伝子を特定した。遺伝子を欠失させると、栄養状態に関わらず常に多細胞体として成長する株が生まれた。
・単細胞増殖している酵母に栄養を与えると、細胞内のMyb1タンパク質が急速に分解され、多細胞体の成長モードへと転換した。逆に、Myb1の発現は単細胞化を誘導した。Myb1が単細胞性―多細胞性変換の分子スイッチであることが示唆された。
・多細胞生物が生まれる仕組みを遺伝子・細胞レベルで研究できる新しいモデル系を確立した。
・海洋由来の黒色酵母注1)株が栄養条件に応じて単細胞性増殖と多細胞体形成を切り換えることを発見した。
・単細胞性―多細胞性の切り換えに必要な10遺伝子を特定した。遺伝子を欠失させると、栄養状態に関わらず常に多細胞体として成長する株が生まれた。
・単細胞増殖している酵母に栄養を与えると、細胞内のMyb1タンパク質が急速に分解され、多細胞体の成長モードへと転換した。逆に、Myb1の発現は単細胞化を誘導した。Myb1が単細胞性―多細胞性変換の分子スイッチであることが示唆された。
・多細胞生物が生まれる仕組みを遺伝子・細胞レベルで研究できる新しいモデル系を確立した。
【研究概要】
名古屋大学大学院理学研究科附属臨海実験所(三重県鳥羽市菅島)の栗田 岳歩 博士後期課程学生と五島 剛太 所長を中心とするグループは、海洋由来酵母における単細胞性―多細胞性転換の細胞内分子基盤を解明しました。
生物進化の過程でいかにして単細胞生物から多細胞生物が生まれたのかはよく分かっていません。一部の生物は環境に応じて単細胞性―多細胞性を切り換える能力を持ちますが、その切り換えを可能にする細胞内基盤を明らかにできれば、多細胞生物誕生の仕組みが明らかになることが期待されていました。
本研究は、海洋由来酵母種が栄養に応じて単細胞性と多細胞性を切り換えることを発見し、遺伝子操作技術の確立を通じて切り換えに必要な遺伝子経路を解明した重要な成果です。地球の生命史の大きな転換点の一つとされる多細胞生物の進化の機構への理解が深まることが期待されます。
本研究成果は、2026年1月8日午前0時(日本時間)付科学雑誌「Nature」に掲載されました。
名古屋大学大学院理学研究科附属臨海実験所(三重県鳥羽市菅島)の栗田 岳歩 博士後期課程学生と五島 剛太 所長を中心とするグループは、海洋由来酵母における単細胞性―多細胞性転換の細胞内分子基盤を解明しました。
生物進化の過程でいかにして単細胞生物から多細胞生物が生まれたのかはよく分かっていません。一部の生物は環境に応じて単細胞性―多細胞性を切り換える能力を持ちますが、その切り換えを可能にする細胞内基盤を明らかにできれば、多細胞生物誕生の仕組みが明らかになることが期待されていました。
本研究は、海洋由来酵母種が栄養に応じて単細胞性と多細胞性を切り換えることを発見し、遺伝子操作技術の確立を通じて切り換えに必要な遺伝子経路を解明した重要な成果です。地球の生命史の大きな転換点の一つとされる多細胞生物の進化の機構への理解が深まることが期待されます。
本研究成果は、2026年1月8日午前0時(日本時間)付科学雑誌「Nature」に掲載されました。
【研究背景と内容】
地球の生命史では、単細胞生物が最初に誕生し、その後、進化の過程で多細胞生物が何度か出現したと考えられています。生物の多細胞化は生命進化史上特筆すべき事象で、動物、植物、菌類など、現在の地球上で見られる複雑な生物の多くは多細胞生物です。しかし、どういった遺伝子レベルの変化が生物の多細胞化を誘導したのか、よく分かっていません。
多細胞生物誕生の仕組みを理解する手がかりとされているのは、同じ遺伝情報を持ちながら環境に応じて単細胞性と多細胞性を切り換える可塑的多細胞生物種です。単細胞性から多細胞性へと切り換えるために必要な遺伝子を特定できれば、多細胞生物誕生の仕組みに迫れると考えられます。
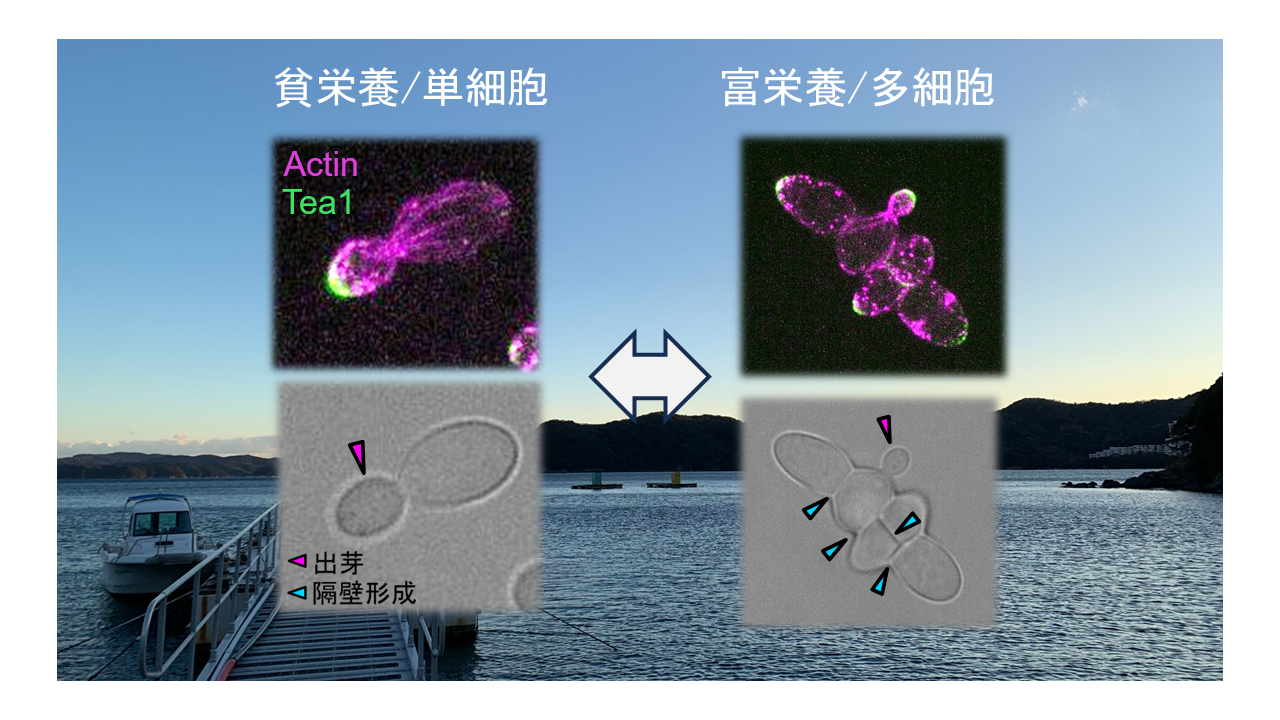
今回の研究では、海洋由来黒色酵母Hortaea werneckii が栄養源依存的に単細胞性増殖と多細胞性成長を切り換えることを発見しました。この酵母は富栄養下では多数の「隔壁注2)形成」を行い多細胞化する一方、貧栄養下では分裂様式を「出芽」に切り替えることで単細胞として増殖しました(図1)。
変異体スクリーニング注3)やライブイメージング注4)を駆使し、この切り換えを可能にする細胞内分子基盤の解明を目指しました。その結果、切り換えに関わる10個の遺伝子を特定しました。興味深いことに、これらの多くは幅広い真菌種において無性胞子形成に関与する遺伝子であり、単細胞/多細胞の切り換えを生み出す遺伝子経路が、祖先生物における既存の遺伝子経路を流用・改変することで進化した可能性が示唆されます。
さらに、機能未知の転写因子注5)Myb1がこの切り換えを制御する極めて重要な遺伝子であることを発見しました。Myb1を細胞から除去させた酵母は単細胞化する能力を失い巨大な多細胞になる一方、Myb1を過剰発現させた酵母は常に単細胞として増殖しました。ライブイメージングの結果、Myb1は貧栄養下では発現量が上昇する一方、栄養を与えるとプロテアソーム注6)依存的に急速に分解されることが判明しました。つまり、Myb1の細胞内タンパク質量は環境中の栄養源の増減と密接に連動して変化しており、これが栄養源依存的な単細胞/多細胞の転換を生み出すスイッチの分子実態であることが解明されました(図2)。
一方、近縁種のNeodothiora pruniでも調べたところ、多くの遺伝子は共通して可塑的多細胞性に関わる一方、興味深いことにMyb1タンパク質にスイッチ機能は認められないことが分かりました。このことは単細胞/多細胞の転換を生み出す分子基盤の柔軟性を示し、Myb1は片方の系統においてスイッチとしての機能を新規に獲得したことを示します。
さらに海洋中のさまざまな環境からHortaea werneckiii株を単離すると、カイメンなどの底生動物注7)より特異的に、多細胞優勢型の株が採取され、カイメンを浸した慣らし培地は酵母の多細胞化を誘導することが分かりました。カイメンは微生物由来の天然化合物を多く含む富栄養なニッチを構成していることが知られ、海洋環境中において酵母の多細胞化を誘導している可能性があります。これらの酵母は富栄養な環境である底生動物などで多細胞化して定着する一方、貧栄養な環境では単細胞化することで拡散し、単細胞/多細胞の切り換えを通じて環境変化に適応していることが考えられます(図3)。
地球の生命史では、単細胞生物が最初に誕生し、その後、進化の過程で多細胞生物が何度か出現したと考えられています。生物の多細胞化は生命進化史上特筆すべき事象で、動物、植物、菌類など、現在の地球上で見られる複雑な生物の多くは多細胞生物です。しかし、どういった遺伝子レベルの変化が生物の多細胞化を誘導したのか、よく分かっていません。
多細胞生物誕生の仕組みを理解する手がかりとされているのは、同じ遺伝情報を持ちながら環境に応じて単細胞性と多細胞性を切り換える可塑的多細胞生物種です。単細胞性から多細胞性へと切り換えるために必要な遺伝子を特定できれば、多細胞生物誕生の仕組みに迫れると考えられます。
今回の研究では、海洋由来黒色酵母Hortaea werneckii が栄養源依存的に単細胞性増殖と多細胞性成長を切り換えることを発見しました。この酵母は富栄養下では多数の「隔壁注2)形成」を行い多細胞化する一方、貧栄養下では分裂様式を「出芽」に切り替えることで単細胞として増殖しました(図1)。
変異体スクリーニング注3)やライブイメージング注4)を駆使し、この切り換えを可能にする細胞内分子基盤の解明を目指しました。その結果、切り換えに関わる10個の遺伝子を特定しました。興味深いことに、これらの多くは幅広い真菌種において無性胞子形成に関与する遺伝子であり、単細胞/多細胞の切り換えを生み出す遺伝子経路が、祖先生物における既存の遺伝子経路を流用・改変することで進化した可能性が示唆されます。
さらに、機能未知の転写因子注5)Myb1がこの切り換えを制御する極めて重要な遺伝子であることを発見しました。Myb1を細胞から除去させた酵母は単細胞化する能力を失い巨大な多細胞になる一方、Myb1を過剰発現させた酵母は常に単細胞として増殖しました。ライブイメージングの結果、Myb1は貧栄養下では発現量が上昇する一方、栄養を与えるとプロテアソーム注6)依存的に急速に分解されることが判明しました。つまり、Myb1の細胞内タンパク質量は環境中の栄養源の増減と密接に連動して変化しており、これが栄養源依存的な単細胞/多細胞の転換を生み出すスイッチの分子実態であることが解明されました(図2)。
一方、近縁種のNeodothiora pruniでも調べたところ、多くの遺伝子は共通して可塑的多細胞性に関わる一方、興味深いことにMyb1タンパク質にスイッチ機能は認められないことが分かりました。このことは単細胞/多細胞の転換を生み出す分子基盤の柔軟性を示し、Myb1は片方の系統においてスイッチとしての機能を新規に獲得したことを示します。
さらに海洋中のさまざまな環境からHortaea werneckiii株を単離すると、カイメンなどの底生動物注7)より特異的に、多細胞優勢型の株が採取され、カイメンを浸した慣らし培地は酵母の多細胞化を誘導することが分かりました。カイメンは微生物由来の天然化合物を多く含む富栄養なニッチを構成していることが知られ、海洋環境中において酵母の多細胞化を誘導している可能性があります。これらの酵母は富栄養な環境である底生動物などで多細胞化して定着する一方、貧栄養な環境では単細胞化することで拡散し、単細胞/多細胞の切り換えを通じて環境変化に適応していることが考えられます(図3)。
【成果の意義】
本研究は、海洋由来酵母を新しい実験モデル系として確立した上で、環境依存的な単細胞/多細胞切り換えの詳細な分子基盤を解明した重要な成果です。切り換えを制御する遺伝子経路の進化の軌跡や自然界における生物学的意義まで、幅広い分野を網羅する統合的な研究です。多細胞性を生み出す進化の機構の理解を得るための手掛かりとなることが期待されます。
本研究は、海洋由来酵母を新しい実験モデル系として確立した上で、環境依存的な単細胞/多細胞切り換えの詳細な分子基盤を解明した重要な成果です。切り換えを制御する遺伝子経路の進化の軌跡や自然界における生物学的意義まで、幅広い分野を網羅する統合的な研究です。多細胞性を生み出す進化の機構の理解を得るための手掛かりとなることが期待されます。
【用語説明】
注1)黒色酵母:
黒色のコロニーを形成する酵母の総称 (真菌類)。
注2)隔壁:
核分裂後、姉妹細胞を分けるのに作られる細胞壁。
注3)変異体スクリーニング:
さまざまな変異株の中から目的の性質を示す株を探索し、その原因遺伝子を明らかにする方法。
注4)ライブイメージング:
細胞を生きたままの状態で顕微鏡観察する方法。
注5)転写因子:
遺伝子発現を制御する機能をもつタンパク質の総称。
注6)プロテアソーム:
細胞内でタンパク分解を担う複合体酵素。シグナル伝達やタンパク質の品質管理などに関わる。
注7)底生動物:
海や湖の底で生活する動物。カイメンやサンゴなど。
注1)黒色酵母:
黒色のコロニーを形成する酵母の総称 (真菌類)。
注2)隔壁:
核分裂後、姉妹細胞を分けるのに作られる細胞壁。
注3)変異体スクリーニング:
さまざまな変異株の中から目的の性質を示す株を探索し、その原因遺伝子を明らかにする方法。
注4)ライブイメージング:
細胞を生きたままの状態で顕微鏡観察する方法。
注5)転写因子:
遺伝子発現を制御する機能をもつタンパク質の総称。
注6)プロテアソーム:
細胞内でタンパク分解を担う複合体酵素。シグナル伝達やタンパク質の品質管理などに関わる。
注7)底生動物:
海や湖の底で生活する動物。カイメンやサンゴなど。
【論文情報】
雑誌名: Nature
論文タイトル:Genetic switch between unicellularity and multicellularity in marine yeasts
著者: Gakuho Kurita, Kyoka A. Adachi, Kazuma Uesaka, Gohta Goshima
DOI : 10.1038/s41586-025-09881-4
URL : https://www.nature.com/articles/s41586-025-09881-4
本件に関するお問い合わせ先
名古屋大学総務部広報課
- TEL
- 052-558-9735
- FAX
- 052-788-6272
- nu_research@t.mail.nagoya-u.ac.jp